产品资料 – 糖生物学与蛋白质工具 – 噬菌体展示
抗 M13 pIII 单克隆抗体
#E8033S 0.1 ml
概述
抗 M13 pIII 单克隆抗体(鼠 IgG2a 同型物)来源于鼠骨髓瘤细胞和脾细胞融合产生的 A23 杂交瘤。BALB/c 小鼠是用 M13 衣壳蛋白 III C 端一半的肽段(259-406 残基)进行免疫。
注意事项
不建议用此抗体做 ELISA 来鉴定完整噬菌体,因为 M13 噬菌体上的抗原决定簇不易暴露。




本产品可直接替代 NEB #E8110S,Ph.D.™-12 噬菌体展示肽库试剂盒
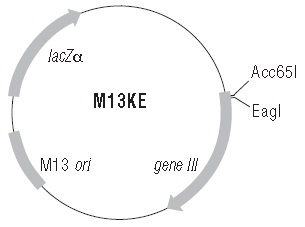
Ph.D.-12™ 噬菌体展示肽库试剂盒 v2 包含 Ph.D.-12 噬菌体展示肽库、淘选实验中用作对照的DYKDDDDK 小鼠单克隆抗体、Protein G 磁珠,以及足够 >50 个测序反应的 -96 gIII 测序引物。Ph.D.-12 噬菌体展示肽库是一种随机 12 肽组合库,随机多肽与 M13 噬菌体的次要外壳蛋白(pIII)的 N 端融合表达。该肽库由约 109 种电转化(即独特的)的序列组成。
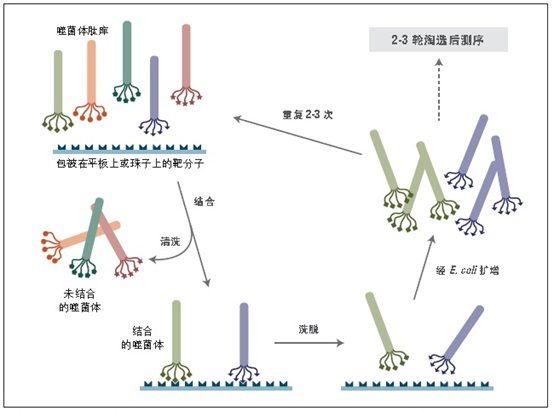
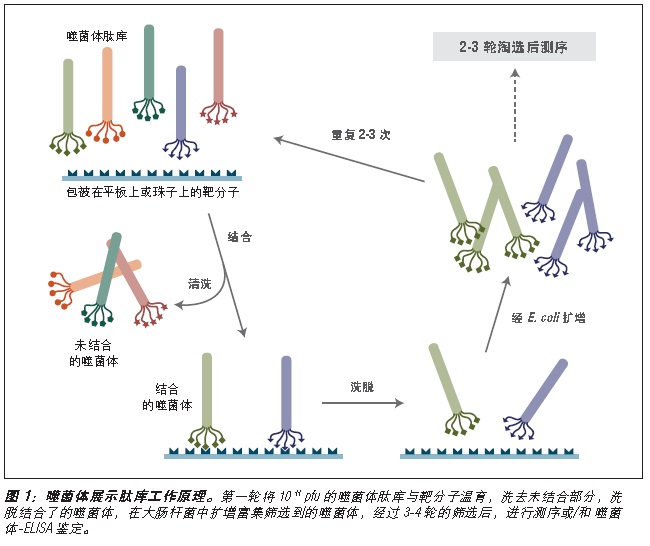
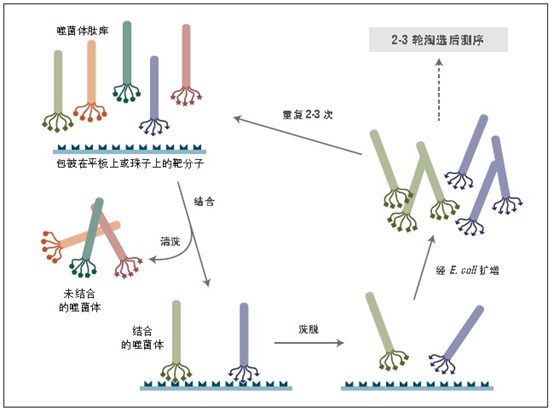
噬菌体展示是一种体外筛选技术。它可以将一系列多肽或蛋白质库展示在噬菌体颗粒的外部,而编码这些蛋白的 DNA 则位于病毒颗粒内部 (1-3)。利用展示蛋白与其编码 DNA 之间的关联,我们可以通过一种称为“生物淘选”的体外筛选方法 (4),快速筛选出与给定靶分子(抗体、酶、细胞表面受体等)亲和结合的噬菌体展示肽。最简单的淘选方法是(见图 1):将展示不同多肽的噬菌体库与固定在平板或小珠上的靶分子孵育,先洗去未结合的噬菌体,而后洗脱特异性结合的噬菌体,扩增洗脱下的噬菌体。重复上述结合/扩增过程,使与靶分子结合的多肽序列的噬菌体得到富集。经过 3-4轮筛选后,每个克隆通过 DNA 测序或 ELISA 鉴定克隆。
本产品可直接替代 NEB # E8100S,Ph.D.™-7 噬菌体展示肽库试剂盒
Ph.D.-7™ 噬菌体展示肽库试剂盒 v2 包含 Ph.D.-7 噬菌体展示肽库、淘选实验中用作对照的DYKDDDDK 小鼠单克隆抗体、Protein G 磁珠,以及足够 >50 个测序反应的 -96 gIII 测序引物。Ph.D.-7 噬菌体展示肽库是一种随机 7 肽组合库,随机多肽与 M13 噬菌体的次要外壳蛋白(pIII)的 N 端融合表达。该肽库由约 109 种电转化(即独特的)的序列组成。